

Психологическая библиотека
ВВЕДЕНИЕ В ПСИХОЛОГИЮ.Учебник для студентов университетов
М., 1999.
Часть II. Биологические процессы и развитие
Глава 2. Нейробиологические основы психологии
Всякое поведение — от моргания глазом до игры в баскетбол и написания компьютерных программ — зависит от интеграции действия множества различных процессов в организме. Такая интеграция обеспечивается нервной системой при поддержке эндокринной системы. Рассмотрим для примера совокупность всех процессов, которые надо эффективно скоординировать, чтобы остановить автомобиль при красном свете светофора. Сначала вам надо увидеть красный свет; это значит, что свет должен быть запечатлен одним из ваших органов чувств — глазами. Нервные импульсы от глаз передаются в мозг, где стимул [В психологии под стимулом обычно понимают предмет или его свойство, воздействующее на орган (или органы) чувств. — Прим. ред.] анализируется и сравнивается с информацией о прошлых событиях, хранящейся в памяти; тогда вы понимаете, что в данном контексте красный свет означает «стоп». Процесс передвижения ноги к педали тормоза и ее нажатия инициируется моторными зонами мозга, контролирующими мышцы ноги и ступни. Чтобы посылать этим мышцам нужные сигналы, мозг должен знать, где находится ступня и куда вы хотите ее переместить. Мозг регистрирует относительные положения частей тела и использует эти данные для построения целенаправленных движений. Однако вы не останавливаете машину одним резким движением ноги. Специальный участок мозга получает непрерывную обратную связь от мышц ноги и ступни, так что вы осознаете величину оказываемого давления на педаль и можете соответственно изменить свои движения. В то же время глаза и некоторые другие органы чувств сообщают, насколько быстро машина останавливается. Если красный свет включился, когда вы мчались к перекрестку, то могут также активироваться некоторые эндокринные железы, что вызовет увеличение частоты сердечных сокращений, учащение дыхания и другие метаболические изменения, связанные с чувством опасности; эти процессы ускоряют ваши реакции в аварийных ситуациях. Ваша остановка на красный свет происходит быстро и кажется автоматической, тем не менее в ней содержится целый ряд сложных сообщений и происходят различные регулировки. Информация, необходимая для осуществления такого рода активности, передается по большим сетям нервных клеток.
Нервная система, органы чувств, мышцы и железы позволяют нам осознавать окружающий мир и приспосабливаться к нему. Восприятие событий зависит от того, как наши органы чувств обнаруживают стимулы и как информация от них интерпретируется мозгом. Поведение человека во многом мотивируется такими потребностями, как голод, жажда и избегание усталости и боли. Способность человека пользоваться речью, мыслить и решать проблемы зависит от работы мозга, который невероятно сложен. Действительно, основу сложнейших мыслительных процессов составляют определенные совокупности электрических и химических явлений в мозге.
В сущности, любые аспекты поведения и психического функционирования легче понять, зная о том, какие биологические процессы лежат в их основе. При рассмотрении восприятия, мотивации и речи в различных частях этой книги мы будем более подробно останавливаться на нейробиологических механизмах. В задачу настоящей главы не входит подробный обзор взаимоотношений биологии и психологии; мы лишь предварительно ознакомимся с некоторыми основными идеями нейробиологии, которые позднее, при обсуждении различных психологических явлений, будут представлены более развернуто.
Нейроны - строительные блоки нервной системы
Основной единицей нервной системы является нейрон — специализированная клетка, передающая нервные импульсы или сигналы другим нейронам, железам и мышцам. Понимать работу нейронов важно потому, что, без сомнения, именно в них таятся секреты функционирования мозга и, соответственно, секреты человеческого сознания. Нам известна их роль в передаче нервных импульсов, и мы знаем, как работают некоторые нервные механизмы; но мы только начинаем узнавать об их более сложных функциях в процессах памяти, эмоций и мышления.
В нервной системе существует два типа нейронов: очень мелкие нейроны, известные как локальные нейроны, и более крупные нейроны, называемые макронейронами. Хотя большинство нейронов являются локальными, мы лишь недавно начали понимать, как они функционируют. Фактически на протяжении долгого времени многие исследователи полагали, что эти крохотные нейроны вовсе не являются нейронами или что они являются незрелыми и неспособными к передаче информации. Сегодня мы знаем, что на самом деле локальные нейроны передают сигналы другим нейронам. Однако они обмениваются сигналами преимущественно с соседними нейронами и не передают информацию на большие расстояния в пределах организма, как это делают макронейроны.
С другой стороны, макронейроны были детально изучены, и поэтому наше внимание будет сосредоточено на этих нейронах. Хотя макронейроны значительно различаются по своим размерам и внешнему виду, все они обладают некоторыми общими характеристиками (см. рис. 2.1) От тела клетки отходит множество коротких отростков, называемых дендритами (от греческого дендрон — дерево). К дендритам и телу клетки поступают нервные импульсы от соседних нейронов. Эти сообщения передаются другим нейронам (или мышцам и железам) через тонкое трубчатое удлинение клетки, которое называется аксоном. Окончание аксона делится на ряд тонких веточек, разветвлений, на концах которых имеются небольшие утолщения, называемые синаптическими окончаниями.
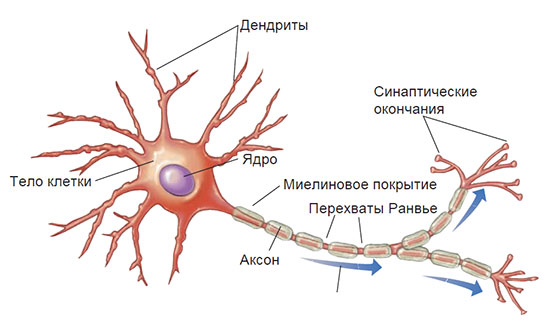
Рис. 2.1. Схематическое строение нейрона.
Стрелками показано направление движения нервного импульса. Некоторые аксоны разветвляются. Эти ответвления называются коллатералями. Аксоны многих нейронов покрыты изолирующей миелиновой оболочкой, что позволяет увеличить скорость передачи нервного импульса.
На самом деле синаптическое окончание не касается возбуждаемого им нейрона. Между синаптическим окончанием и телом или дендритом воспринимающей клетки существует небольшой промежуток. Такое сопряжение называется синапсом, а сам промежуток называется синаптической щелью. Когда нервный импульс, проходя по аксону, достигает синаптического окончания, он запускает выделение химического вещества, называемого нейромедиатором (или просто медиатором). Медиатор проникает через синаптическую щель и стимулирует следующий нейрон, передавая тем самым сигнал от одного нейрона к другому. Аксоны от очень многих нейронов синаптически контактируют с дендритами и телом клетки отдельного нейрона (рис. 2.2).
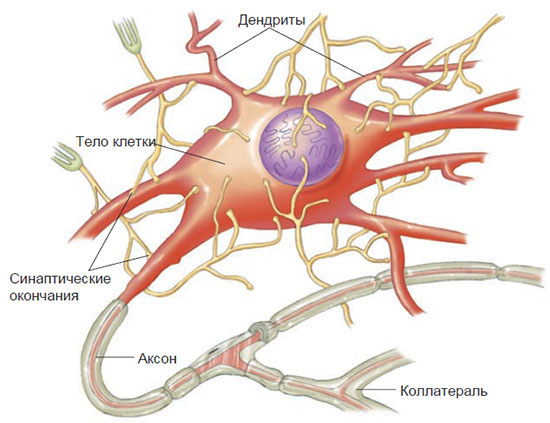
Рис. 2.2. Синапсы на клеточном теле нейрона.
Множество различных аксонов, каждый из которых многократно разветвляется, синаптически контактируют с дендритами и телом клетки отдельного нейрона. Каждое концевое ответвление аксона имеет утолщение, которое называется синаптическим окончанием и содержит химическое вещество, высвобождаемое и передаваемое нервным импульсом через синапс к дендритом или телу клетки воспринимающего нейрона.
Хотя все нейроны обладают этими общими признаками, они весьма разнообразны по форме и величине (рис. 2.3). У нейрона спинного мозга аксон может достигать 3-4 футов длины и идти от конца позвоночника до мышц большого пальца ступни; нейрон головного мозга может иметь размер всего лишь в несколько тысячных долей дюйма.
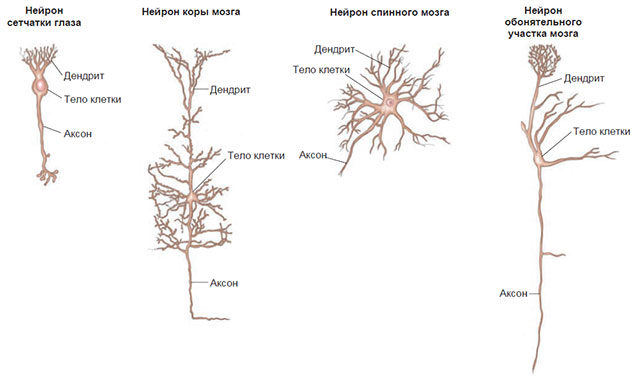
Рис. 2.3. Формы и относительные величины нейронов.
Аксон нейрона спинного мозга может достигать нескольких футов длины (на рисунке показан не полностью).
В зависимости от выполняемых ими общих функций нейроны делятся на три категории. Сенсорные нейроны передают импульсы от рецепторов в центральную нервную систему. Рецепторы — это специализированные клетки органов чувств, мышц, кожи и суставов, способные обнаруживать физические или химические изменения и преобразовывать их в импульсы, проходящие по сенсорным нейронам. Моторные нейроны несут сигналы, выходящие из головного или спинного мозга, к исполнительным органам, т. е. к мышцам и железам. Промежуточные нейроны получают сигналы от сенсорных нейронов и посылают импульсы к другим промежуточным нейронам и к моторным нейронам. Промежуточные нейроны обнаружены только в головном мозге, глазах и спинном мозге.
Нерв — это пучок длинных аксонов, принадлежащих сотням или тысячам нейронов. Один нерв может содержать аксоны как от сенсорных, так и от моторных нейронов.
Помимо нейронов в нервной системе есть множество клеток, не являющихся нервными, но рассеянных между — и часто вокруг — нейронов; их называют глиальными клетками. Количество глиальных клеток превосходит число нейронов в 9 раз, и они занимают больше половины объема мозга. Их название (от греческого glia — клей) определяется одной из их функций — закреплением нейронов на их местах. Кроме того, они вырабатывают питательные вещества, необходимые для здоровья нейронов, и как бы «ведут хозяйство», очищая нейрональную среду (на синаптических участках), тем самым поддерживая сигнальную способность нейронов. Бесконтрольное разрастание глиальных клеток — причина почти всех опухолей мозга.
Оценки количества нейронов и глиальных клеток в нервной системе человека широко варьируются и зависят от метода подсчета; пока ученые не пришли к единому мнению об их количестве. Только в самом мозге человека, по разным оценкам, насчитывается от 10 миллиардов до 1 триллиона нейронов; независимо от предполагаемого количества нейронов количество глиальных клеток примерно в 9 раз больше (Groves & Rebec, 1992). Эти цифры кажутся астрономическими, но такое количество клеток бесспорно необходимо, учитывая всю сложность поведения человека.
Потенциалы действия
Информация передается по нейрону в виде нейронного импульса, называемого потенциалом действия — электрохимическим импульсом, проходящим от дендритовой области к окончанию аксона. Каждый потенциал действия является результатом движения электрически заряженных молекул, называемых ионами, осуществляемого внутри и снаружи нейрона. Описанные ниже электрические и химические процессы приводят к формированию потенциала действия.
Клеточная мембрана является полупроницаемой; это означает, что некоторые химические вещества могут легко проходить через клеточную мембрану, тогда как другие не пропускаются через нее, за исключением тех случаев, когда специальные проходы в мембране открыты. Ионные каналы — это белковые молекулы наподобие пончиков, образующие поры в клеточной мембране (рис. 2.4). Открывая или закрывая поры, эти белковые структуры регулируют поток электрически заряженных ионов, таких как натрий (Na+), калий (K+), кальций (Са++) или хлор (Сl-). Каждый ионный канал действует избирательно: когда он открыт, то пропускает через себя только один тип ионов.
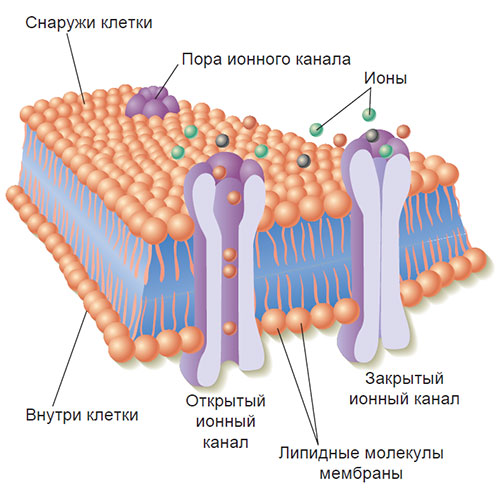
Рис. 2.4. Ионные каналы.
Такие химические вещества, как натрий, калий, кальций и хлор, проходят сквозь клеточную мембрану через торообразные протеиновые молекулы, называемые ионными каналами.
Нейрон, когда он не передает информацию, называют покоящимся нейроном. В покоящемся нейроне отдельные протеиновые структуры, называемые ионными насосами, помогают поддерживать неравномерное распределение различных ионов по клеточной мембране путем перекачивания их внутрь или вне клетки. Например, ионные насосы транспортируют Na+ за пределы нейрона каждый раз, когда он проникает в нейрон, и закачивают K+ обратно в нейрон каждый раз, когда он выходит наружу. Таким образом, у нейрона в состоянии покоя поддерживается высокая концентрация Na+ снаружи и низкая концентрация внутри клетки. Действие этих ионных каналов и насосов создает поляризацию клеточной мембраны, которая имеет положительный заряд с наружной и отрицательный заряд с внутренней стороны.
Когда нейрон, находящийся в состоянии покоя, стимулируется, разность потенциалов на клеточной мембране уменьшается. Если падение напряжения достаточное, натриевые каналы в точке стимуляции на короткое время открываются и ионы Na+ проникают внутрь клетки. Этот процесс называется деполяризацией; теперь внутренняя сторона мембраны в этом участке оказывается заряженной положительно относительно внешней. Соседние натриевые каналы чувствуют это падение напряжения и в свою очередь открываются, вызывая деполяризацию прилежащих участков. Такой самоподдерживаемый процесс деполяризации, распространяющейся вдоль тела клетки, называется нервным импульсом. По мере продвижения этого импульса по нейрону натриевые каналы за ним закрываются и включаются ионные насосы, быстро восстанавливающие в клеточной мембране исходное состояние покоя (рис. 2.5).
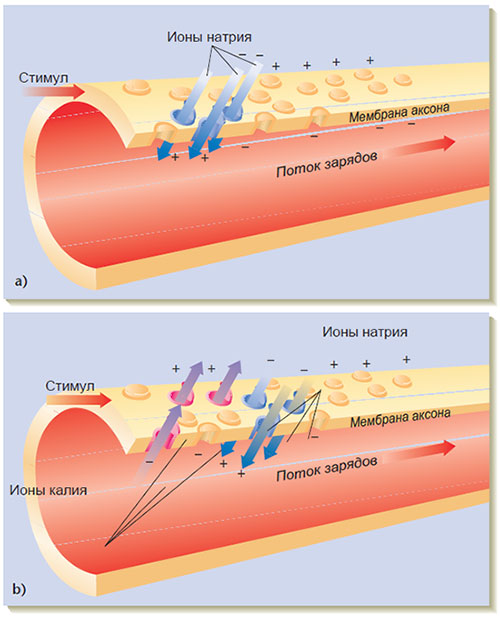
Рис. 2.5. Потенциал действия.
а) В течение действия потенциала натриевые шлюзы в мембране нейрона открыты и ионы натрия входят внутрь аксона, неся с собой положительный заряд, б) Когда потенциал действия возникает в какой-либо точке аксона, натриевые шлюзы закрываются в этой точке и открываются в следующей, расположенной по длине аксона. Когда натриевые шлюзы закрыты, открыты калиевые шлюзы и ионы калия выходят из аксона, унося с собой положительный заряд (по материалам Starr & Taggart, 1989).
Скорость продвижения нервного импульса по аксону может меняться от 3 до 300 км/час, в зависимости от диаметра аксона: как правило, чем больше диаметр, тем выше скорость. Скорость может зависеть также от того, есть ли у аксона миелиновое покрытие. Это покрытие состоит из специальных глиальных клеток, окутывающих аксон и идущих одна за другой с небольшими перехватами (промежутками) (как на рис. 2.1). Эти маленькие промежутки называют узлами Ранвьера. Благодаря изолирующим свойствам миелинового покрытия нервный импульс как бы прыгает от одного узла Ранвьера к другому — процесс, известный как салтаторная проводимость, что значительно повышает скорость передачи по аксону. (Термин салтаторная происходит от латинского слова saltare, что означает «прыгать».) Наличие миелиновых покрытий характерно для высших животных и особенно широко распространено в тех частях нервной системы, где скорость передачи — решающий фактор. Рассеянный склероз, сопровождаемый серьезными сенсомоторными дисфункциями нервной системы, — это заболевание, при котором организм разрушает свой собственный миелин.
Синаптическая передача импульсов
Синаптическое сопряжение между нейронами чрезвычайно важно, поскольку именно здесь клетки передают свои сигналы. Отдельный нейрон разряжается или возбуждается, когда приходящая к нему через множество синапсов стимуляция превышает определенный порог. Нейрон разряжается одним коротким импульсом и затем несколько тысячных долей секунды остается инактивным. Величина нервного импульса постоянна, и он не может быть вызван до тех пор, пока стимул не достигнет порогового уровня; это называется законом «все или ничего». Нервный импульс, раз начавшись, распространяется по аксону, достигая множества его окончаний.
Как мы уже говорили, в синапсе нейроны не контактируют непосредственно; здесь есть небольшая щель, через которую сигнал и должен быть передан (рис. 2.6). Когда нервный импульс продвигается по аксону и достигает синаптического окончания, он стимулирует находящиеся там синаптические пузырьки. Они представляют собой маленькие шарики, в которых содержатся нейротрансмиттеры; при стимуляции пузырьки выпускают эти нейротрансмиттеры. Нейротрансмиттеры проникают через синаптическую щель-зазор и захватываются молекулами воспринимающего нейрона, находящимися в его клеточной мембране. Молекулы медиатора и рецептора подходят друг к другу примерно так, как кусочки разрезной головоломки или ключ к замку. На основе соотношения двух молекул по принципу «ключ—замок» изменяется проницаемость мембраны воспринимающего нейрона. Некоторые медиаторы, находящиеся в связке со своими рецепторами, оказывают возбуждающее действие и увеличивают проницаемость в сторону деполяризации, а некоторые оказывают тормозящее действие и уменьшают проницаемость. При возбуждающем действии вероятность возбуждения нейрона увеличивается, а при тормозящем — уменьшается.
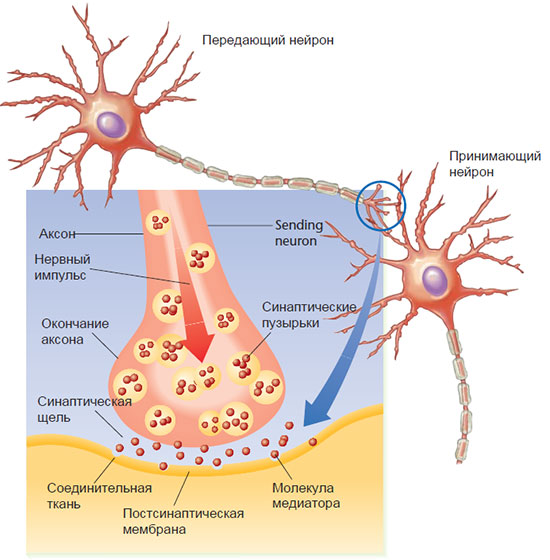
Рис. 2.6. Высвобождение медиаторов в синаптическую щель.
Медиатор доставляется к пресинаптической мембране в синаптических пузырьках, которые смешиваются с этой мембраной, высвобождая свое содержимое в синаптическую щель. Молекулы медиатора проникают через щель и соединяются с рецепторными молекулами постсинаптической мембраны.
Один нейрон может иметь многие тысячи синапсов с сетью других нейронов. Некоторые из этих нейронов высвобождают возбуждающие медиаторы, другие — тормозящие. В зависимости от характерного для них паттерна передачи импульсов (firing) различные аксоны высвобождают различные вещества-медиаторы в разное время. Если в определенное время и на определенном участке клеточной мембраны возбуждающие воздействия на воспринимающий нейрон начинают превышать тормозящие, то происходит деполяризация и нейрон разряжается импульсом соответственно закону «все или ничего».
Электронная микрофотография нейрона, плотно упакованного синапсами
После высвобождения молекул медиатора и прохождения их через синаптическую щель их действие должно быть очень коротким. В противном случае воздействие медиатора будет длиться слишком долго и точный контроль станет невозможным. Кратковременность действия достигается одним из двух путей. Некоторые медиаторы почти мгновенно удаляются из синапса посредством обратного захвата — процесса, при котором медиатор снова поглощается синаптическими окончаниями, откуда он был выпущен. Обратный захват прекращает действие медиатора и избавляет окончания аксона от необходимости дополнительно производить это вещество. Действие других медиаторов прекращается благодаря деградации — процессу, при котором ферменты, содержащиеся в мембране воспринимающего нейрона, инактивируют медиатор, химически разрушая его.
Нейротрансмиттеры
Известно более 70 различных медиаторов, и нет сомнений, что будут открыты еще. Помимо этого, некоторые медиаторы могут связываться более чем с одним типом рецепторных молекул и вызывать при этом различные эффекты. Например, нейротрансмиттер глутамат может активизировать как минимум 16 различных типов рецепторных молекул, позволяя нейронам реагировать различным образом на этот один и тот же нейротрансмиттер (Westbrook, 1994). Некоторые нейротрансмиттеры являются возбуждающими в одних зонах и тормозящими в других, так как в этих процессах участвуют два различных типа рецепторных молекул. В этой главе мы, конечно, не сможем рассказать о всех нейротрансмиттерах, обнаруженных в нервной системе, поэтому подробно остановимся на некоторых из них, оказывающих существенное влияние на поведение.
Ацетилхолин (АЦХ) обнаружен во многих синапсах по всей нервной системе. Вообще, это возбуждающий нейротрансмиттер, но он может быть и тормозящим, в зависимости от того, какой тип молекулы рецептора находится в мембране воспринимающего нейрона. Особенно часто АЦХ встречается в гиппокампе — зоне переднего мозга, играющей ключевую роль в формировании новых следов памяти (Squire, 1987).
Болезнь Альцгеймера (предстарческий склероз мозга. — Прим. перев.) — тяжелое нарушение, часто встречающееся в пожилом возрасте и сопровождающееся нарушениями памяти и других когнитивных функций. Было показано, что при болезни Альцгеймера вырождаются нейроны переднего мозга, производящие АЦХ, и соответственно снижается способность мозга производить АЦХ; чем меньше АЦХ производится передним мозгом, тем обширнее потеря памяти.
АЦХ выделяется также во всех синапсах, образованных между нервными окончаниями и волокнами скелетной мускулатуры. АЦХ подводится к концевым пластинкам — небольшим образованиям, расположенным на клетках мышц. Концевые пластинки покрыты молекулами рецептора, которые при активации их ацетилхолином запускают химическую реакцию между молекулами внутри мышечных клеток, заставляя их сокращаться. Некоторые препараты, влияющие на АЦХ, могут вызывать паралич мышц. Например, яд ботулин, выделяемый некоторыми видами бактерий в плохо закрытых консервах, блокирует выделение АЦХ в нервно-мышечных синапсах и может вызвать смерть от паралича дыхательных мышц. Некоторые нервные газы военного назначения, а также многие пестициды вызывают паралич путем разрушения ферментов, расщепляющих АЦХ после включения нейрона; когда процесс расщепления нарушен, в нервной системе происходит неконтролируемое накопление АЦХ и нормальная синаптическая передача становится невозможной.
Норэпинефрин (НЭ) — это медиатор, продуцируемый многими нейронами ствола мозга. Такие хорошо известные препараты, как кокаин и амфетамины, продлевают действие норэпинефрина путем замедления его обратного захвата. Из-за задержки обратного захвата воспринимающий нейрон активируется дольше, чем и объясняется психостимулирующий эффект этих препаратов. Литий, наоборот, ускоряет обратный захват НЭ, вызывая у человека подавленное настроение. Всякое вещество, повышающее или понижающее уровень НЭ в мозге, соответственно повышает или снижает настроение человека.
Допамин. Химически допамин очень близок к норэпинефрину. Высвобождение допамина в определенных зонах головного мозга вызывает интенсивное ощущение удовольствия, и в настоящий момент проводятся исследования, изучающие роль допамина в развитии пристрастий. Избыток допамина в определенных зонах мозга может вызывать шизофрению, тогда как его недостаток в других зонах может приводить к болезни Паркинсона. Лекарства, используемые для лечения шизофрении, например торазин или клозапин, блокируют рецепторы допамина. В противовес им препарат L-dopa, чаще всего прописываемый страдающим болезнью Паркинсона, увеличивает количество допамина в мозге.
Серотонин. Серотонин принадлежит к той же группе химических препаратов, называемых моноаминами, что и допамин и норэпинефрин. Как и норэпинефрин, серотонин играет важную роль в регулировании настроения. Так, низкий уровень серотонина ассоциируется с ощущением депрессии. Были разработаны специфические антидепрессанты, называемые селективными ингибиторами обратного захвата серотонина (СИОЗС), повышающие уровень серотонина в мозге путем блокирования обратного захвата серотонина пресинаптическими окончаниями нейронов. Прозак, Золофт и Паксил, лекарственные препараты, как правило прописываемые для лечения депрессии, — являются ингибиторами обратного захвата серотонина. Серотонин также играет важную роль в регуляции сна и аппетита, а потому используется также при лечении расстройства питания — булимии. Изменяющий настроение препарат ЛСД оказывает свое воздействие, повышая уровень серотонина в мозге. ЛСД по своему химическому строению похож на медиатор серотонин. влияющий на эмоции. Данные показывают, что ЛСД накапливается в некоторых клетках мозга, где имитирует действие серотонина и тем самым создает повышенную стимуляцию этих клеток.
ГАМК. Еще один широкоизвестный медиатор — гамма-аминомасляная кислота (ГАМК), являющаяся одним из основных тормозных медиаторов в нервной системе. Например, препарат пикротоксин блокирует рецепторы ГАМК и вызывает конвульсии, поскольку из-за недостатка тормозного действия ГАМК контроль за движением мышц становится затрудненным. Некоторые транквилизаторы, основанные на свойстве ГАМК усиливать торможение, применяются для лечения пациентов, страдающих тревожностью.
Глутамат. Возбуждающий медиатор глутамат присутствует в большем количестве нейронов центральной нервной системы, чем любой другой медиатор. Существует как минимум три подтипа глутаматовых рецепторов, и один из них, как полагают, играет роль в научении и памяти. Он называется рецептором НМДА — по названию вещества, применяемого для его обнаружения (N-метил D-аспартат). Больше всего НМДА-рецепторов содержится в нейронах гиппокампа (участка около середины мозга), и есть различные данные, показывающие, что эта зона играет решающую роль в формировании новых следов памяти.
Рецепторы НМДА отличаются от других рецепторов тем, что для их активации нужны последовательные сигналы от двух различных нейронов. Сигнал от первого из них повышает чувствительность клеточной мембраны, в которой находится рецептор НМДА. После повышения чувствительности второй сигнал (глутаминовый медиатор от другого нейрона) сможет активировать этот рецептор. При получении такого сдвоенного сигнала рецептор НМДА пропускает в нейрон очень много ионов кальция. Их приток вызывает долговременное изменение в мембране нейрона, делая ее более чувствительной к первоначальному сигналу, когда тот повторится в следующий раз; это явление называют долговременной потенциацией, или ДП (рис. 2.7).
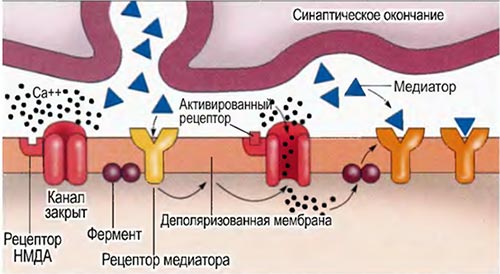
Рис. 2.7. Рецепторы НМДА и долговременная потенциация.
На схеме показан возможный механизм влияния рецепторов НМДА на долговременное изменение силы синаптической связи (эффект ДП). Когда первый передающий нейрон высвобождает медиаторы, они активируют не-НМДА рецепторы воспринимающего нейрона (1), которые частично деполяризуют клеточную мембрану (2). Эта частичная деполяризация повышает чувствительность НМДА-рецепторов, так что теперь их могут активировать глутаматовые медиаторы, высвобождаемые вторым передающим нейроном (3). Активация НМДА-рецепторов заставляет открыться связанные с ними кальциевые каналы (4). Ионы кальция поступают в клетку и взаимодействуют с различными ферментами (5), что, как полагают, приводит к перестройке клеточной мембраны (6). В результате перестройки у воспринимающего нейрона повышается чувствительность к медиаторам, высвобождаемым первым нейроном, так что последний со временем сможет сам по себе активировать воспринимающий нейрон; так возникает эффект долговременной потенциации.
Такой механизм, в котором два конвергирующих сигнала усиливают синаптическую связь, может объяснить, как отдельные события ассоциируются в памяти. Например, в эксперименте с ассоциативным научением вслед за звуком колокольчика немедленно показывалась пища. Когда собака видит пищу, у нее выделяется слюна. Но при повторяющемся сочетании звука и пищи собака научается выделять слюну только на звук колокольчика: это может указывать на то, что сигнал «колокольчик» и сигнал «пища» конвергировали на синапсах, вызывающих слюноотделение. При достаточно многократном предъявлении пары «колокольчик—еда» эти синаптические связи усиливаются под влиянием ДП, и со временем один только звук колокольчика заставляет собаку выделять слюну. На основе механизма НМДА создана любопытная теория ассоциирования событий в памяти, которая сейчас активно развивается (Malonow, 1994; Zalutsky & Nicoll, 1990).
Исследования нейротрансмиттеров и рецепторов получили широкое практическое применение. Некоторые из сфер их применения описаны в рубрике «На переднем крае психологических исследований» на следующей странице.


