

Психологическая библиотека
ВВЕДЕНИЕ В ПСИХОЛОГИЮ.Учебник для студентов университетов
М., 1999.
Часть III. Сознание и восприятие
Глава 5. Восприятие
Информация может поступать к органам чувств в виде отдельных единиц и фрагментов, но не таким мы воспринимаем этот мир. Мы воспринимаем мир предметов и людей, воздействующих на нас как единые целостные образования, а не как конгломераты ощущений. Только при необычных обстоятельствах либо во время черчения или рисования мы начинаем замечать отдельные признаки и части стимулов; практически все остальное время мы видим трехмерные объекты и слышим слова или музыку.
Изучая восприятие, мы обращаемся к тому, как сенсорная информация интегрируется в перцепты объектов и как эти перцепты затем используются для ориентировки в окружающем мире (перцепт — это продукт процесса восприятия). Ученые все больше ориентируются на такой подход к исследованию восприятия, в котором ставится вопрос: какие задачи позволяет решать устройство данной перцептивной системы. Постоянно упоминаются две общие задачи. Перцептивная система должна определять: а) что это за объекты (яблоки, столы, кошки или еще что-то) и б) где эти объекты находятся (слева на расстоянии моей вытянутой руки, в сотне метров прямо передо мной или еще где-то). Эти же две задачи стоят перед слуховым восприятием (что это был за звук — телефон или сирена и откуда он пришел — спереди или сбоку) и перед другими сенсорными модальностями.
В случае зрения определение того, что это за объект, обычно связывают с процессом распознавания паттернов, или просто распознаванием. Для выживания оно имеет решающее значение, поскольку прежде чем делать вывод о каких-либо важных свойствах объекта, чаще всего надо сначала знать, что это за объект. Так, когда мы уже знаем, что объект — это яблоко, мы знаем и то, что он съедобен: а когда известно, что объект — это волк, мы знаем, что его лучше не беспокоить. Определение местоположения видимых объектов называется пространственной локализацией, или просто локализацией. Она тоже необходима для выживания. Благодаря локализации мы передвигаемся среди окружения. Если бы такой способности не было, мы бы постоянно наталкивались на объекты, не могли бы взять вещь, к которой тянемся, и натыкались бы на опасные предметы и хищников.
Помимо распознавания и локализации объектов другая задача нашей перцептивной системы состоит в том, чтобы сохранять постоянство видимых объектов, несмотря на то что их отпечатки на сетчатках глаз постоянно меняются. Эта константность восприятия также будет рассмотрена нами.
В ходе дальнейшего обсуждения мы сначала обратимся к выяснению того, как мозг распределяет решаемые им перцептивные задачи. Затем мы обратимся к тому, что на сегодня известно об основных процессах восприятия — локализации, распознавании и константности восприятия. При этом мы также обсудим роль внимания. Наконец, мы рассмотрим развитие восприятия. На протяжении всей этой главы мы прежде всего сосредоточимся на зрительном восприятии, поскольку это наиболее исследованная область. Не забывайте, однако, что задачи локализации, распознавания и константности, по-видимому, имеют отношение ко всем сенсорным модальностям. Возьмем, например, распознавание: с помощью слуха можно распознать сонату Моцарта, с помощью обоняния — жареную еду из Макдональдса, с помощью осязания — монету 25 центов в кармане брюк, а с помощью чувства тела — что танцуешь польку.
Разделение труда в мозге
За последнее десятилетие мы многое узнали о нервных процессах, лежащих в основе восприятия. В самом общем плане можно сказать, что часть мозга, отвечающая за зрение — зрительная кора, — функционирует по принципу разделения труда. Не вся зрительная кора участвует во всех или почти всех аспектах восприятия, а различные области специализированы для выполнения различных перцептивных функций (Kosslyn & Koenig, 1992; Zeki, 1993).
Зрительная кора
Более тысячи миллионов нейронов коры головного мозга восприимчивы к зрительным входным сигналам. Все, что нам известно об этих нейронах и о механизмах их функционирования, мы узнали благодаря использованию лишь небольшого числа техник. Наши знания, полученные в ходе исследований, проводимых на животных, основаны прежде всего на экспериментах, в которых (с помощью микроэлектродов) производилась запись электрических импульсов, идущих от отдельной клетки, о чем рассказывалось в главе 4. Развитие современных техник, используемых при проведении таких исследований, во многом обязано пионерским работам Дэвида Хьюбела и Торнстейна Визела (David Hubel and Tornstein Wiesel), получивших Нобелевскую премию в 1981 году.
Источником большей части наших знаний, полученных в ходе исследований с участием людей, являются «естественные эксперименты», т. е. случаи повреждений мозга у людей, проливающие свет на связь визуального поведения со специфическими участками мозга. В число исследователей, работающих в этой области, входят нейрологи (медицинские работники, специализирующиеся на лечении мозга), нейропсихологи (психологи, специализирующиеся на лечении и изучении пациентов с повреждениями мозга). Прекрасным введением в данную область исследований может послужить книга Оливера Сэка «Человек, который спутал свою жену со шляпой» (Oliver Sack. The Man Who Mistook His Wife for a Hat, 1987).
Наиболее впечатляющие открытия последних лет, касающиеся человеческого мозга, были сделаны с помощью получения изображений мозга без хирургического вмешательства. Эта область, получившая название визуальных исследований мозга, включает такие техники, как связанные с событиями потенциалы (event-related potentials, ERP), позитронно-эмиссионная томография (ПЭТ) и функциональные магнитно-резонансные изображения (ФМРИ).
Наиболее важным участком мозга, ответственным за обработку визуальной информации, является область, называемая первичной зрительной корой, или зоной V1. Она расположена в задней, или затылочной, части мозга, показанной на рис. 5.1. Именно с этим участком коры большого мозга непосредственно соединены нейроны, посылающие зрительные сигналы от глаз. Все остальные чувствительные к визуальным сигналам участки мозга (их было идентифицировано более 30) связаны с глазами через зону V1.
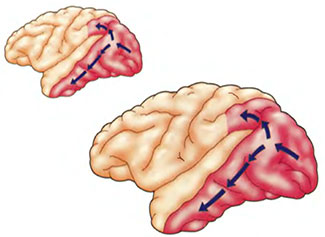
Система локализации показана стрелками, идущими от задней части мозга к верхней; система распознавания показана стрелками от задней части мозга к нижней (по: Mishkin, Ungerleider & Macko, 1983).
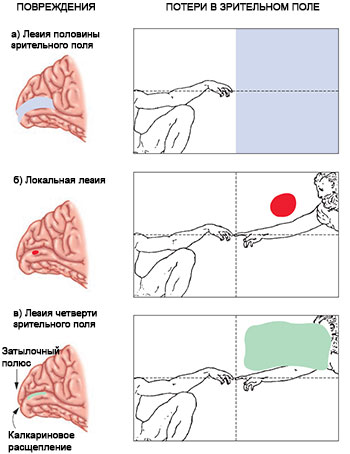
Как это нередко случалось в истории психологии, функции зоны V1 были выяснены задолго до того, как появились современные техники регистрации нейронных сигналов и получения изображений мозга. Впервые эти функции стали очевидными, когда врачи обследовали пациентов, страдающих от локальных повреждений головы, полученных на войне. Была обнаружена связь между повреждениями тканей (технически называемыми лезиями) специфических участков зоны V1 и слепотой в отдельных зонах зрительного поля (технически называемых скотомами) (см. рис. 5.2). Заметьте, что данная форма слепоты не была вызвана поражениями глаз или оптического нерва; она имеет исключительно кортикальное происхождение. Например, самая центральная зона зрительного поля — фовеа — будет страдать от скотомы, если имеет место лезия наиболее удаленного к затылку участка зоны V1. Скотомы более удаленных к периферии зон зрительного поля вызываются лезиями участков, расположенных ближе к передней части зоны V1. Создается впечатление, будто карта зрительного поля натянута на заднюю часть коры, а ее центр приходится на наиболее удаленную заднюю точку коры.
«Карта» зрительного поля перевернута вверх ногами и отражена зеркально.
Однако эта «карта» перевернута вверх ногами и отражена зеркально. Точки, расположенные в верхней половине зрительного поля, нанесены на карту ниже основной кортикальной складки, или долины, а точки, расположенные в нижней половине зрительного поля, представлены на карте выше этой складки. Левая половина зрительного поля соответствует правой стороне зоны V1, а правая половина зрительного поля — ее левой стороне.
Нейроны первичной зрительной коры восприимчивы ко многим характеристикам, которые содержит зрительный образ, таким как яркость, цвет, ориентация в пространстве и движение. Однако одной из наиболее важных характеристик этих нейронов является то, что каждый из них отвечает за анализ лишь очень небольшой области образа. В фовеальной части образа эти области могут быть столь малы, что позволяют рассмотреть детали, размеры которых меньше одного 1 миллиметра, на расстоянии вытянутой руки. Эти нейроны также взаимодействуют между собой только в пределах очень ограниченных областей. Преимущество такой организации состоит в том, что она позволяет одновременно производить очень подробный анализ всего зрительного поля. Однако такой анализ не позволяет координировать информацию о точках зрительного образа, не расположенных в непосредственной близости друг от друга; иными словами, она не позволяет «за деревьями видеть лес».
Для выполнения этой задачи кортикальные нейроны посылают информацию из зоны V1 во многие другие области мозга, анализирующие зрительную информацию. Каждая из этих областей специализируется на выполнении определенной задачи, например, анализе цвета, движения, формы и расположения в пространстве. Эти выполняющие более специфические задачи области также постоянно взаимодействуют с зоной V1, так что нейронную коммуникацию между этими областями правильнее будет представлять себе как переговоры, а не как одностороннее отдавание приказов (Damasio, 1990; Zeki, 1993). Одна из наиболее важных форм разделения труда при визуальном анализе, производимом мозгом, состоит в различии между локализацией и распознаванием, к рассмотрению которого мы и переходим.
Система распознавания и система локализации
Положение о том, что локализация объекта и его распознавание — качественно различные задачи, подтверждается данными, согласно которым эти задачи выполняются различными участками зрительной коры. Распознавание объектов зависит от того отдела зрительной системы, в который входит воспринимающая зона зрительной коры (это первый пункт приема зрительной информации в коре), а также участок, находящийся около нижней части коры полушарий. Локализацией же объектов занимается отдел зрительной системы, в который входит воспринимающая часть зрительной коры, а также участок коры рядом с верхней частью мозга (рис. 5.1). Эксперименты с нечеловекообразными обезьянами показали, что если разрушить отдел распознавания в зрительной системе животного, то оно все еще сможет выполнять задачи, связанные с восприятием пространственных отношений между объектами (например, когда один предмет находится перед другим), но не сможет выполнять задачи, требующие различения реальных предметов, — например, отличать куб от цилиндра; если же разрушить отдел локализации, то животное сможет выполнять задачи, где требуется отличить куб от цилиндра, но не справится с задачей, в которой надо определять взаимное расположение предметов (Mishkin, Ungelleider & Macko, 1983).
В более современных исследованиях для подтверждения существования раздельных систем распознавания и локализации объектов в мозге человека были применены методы компьютерной томографии. Наиболее часто применяется позитронно-эмиссионная томография (ПЭТ), о которой рассказывалось в гл. 2. Вспомним ее основной принцип. Пациенту в кровоток впрыскивается радиоактивный краситель, затем его помещают в ПЭТ-сканер и задают разные задачи. ПЭТ-сканер показывает рост радиоактивности в некоторых участках мозга, что указывает на увеличение кровотока в этих участках. Предполагается, что участки, где кровоток больше, участвуют в выполнении текущей задачи.
В одном из исследований с использованием ПЭТ испытуемым давались две задачи: одна — тест на узнавание лиц, т. е. задача на распознавание, а другая — тест мысленного вращения, который предположительно связан с локализацией. В задаче на узнавание лиц испытуемому в каждой пробе предъявлялась целевая картинка, а под ней — два тестовых изображения лица. На одном из тестовых изображений было то же лицо, что и на целевой картинке, но в измененной ориентации и при другом освещении; на другом — лицо другого человека. Испытуемый должен был решить, на какой из тестовых картинок изображено то же лицо, что и на целевой картинке (рис. 5.3а). При выполнении этой задачи наблюдался рост кровотока в распознающем отделе коры (ветвь, оканчивающаяся у основания коры), чего не наблюдалось в отделе, отвечающем за локализацию (ветвь, оканчивающаяся у вершины коры). В задаче с мысленным вращением результаты были совсем другими. Здесь испытуемым в каждой пробе показывали целевую картинку, на которой была двойная линия, а на некотором расстоянии от нее — точка. Под целевой картинкой были две тестовые. На одной из них была та же конфигурация, что и на цели, но повернутая на определенный угол; на другой была конфигурация с другим расположением точки и линий (рис. 5.3б). При решении этой задачи у испытуемых наблюдался рост кровотока в отделе коры, отвечающем за локализацию, чего не наблюдалось в отделе, отвечающем за распознавание. Следовательно, процессы локализации и распознавания протекают в совершенно различных участках зрительной коры (Grady et al., 1992; Haxby et al., 1990).

Примеры рисунков, предъявлявшихся в задачах на сравнение а) лиц и 6) расположений точки (по: Grady et al., 1992).
Разделение труда в зрительной коре не ограничивается делением между локализацией и распознаванием. Оказывается, что разные виды информации, используемой при распознавании, — например, цвет, форма, текстура — обрабатываются различными участками или клетками распознающего отдела коры.
Аналогичным образом, различным видам информации, используемым при распознавании — форме, цвету и текстуре, — также соответствуют специализированные подотделы мозга, ответственные за их анализ (Livingstone & Hubel, 1988; Zeki, 1993). Таким образом, зрительная кора состоит из многочисленных «модулей переработки», каждый из которых специализируется на выполнении определенной задачи. Чем больше мы узнаем о нейрологическом базисе других сенсорных модальностей (а также о других психологических функциях), тем более правдоподобной представляется данная модель, предполагающая разделение труда между различными модулями переработки информации.


